
封面报道
我们为何会变老?
生命时钟:端粒磨损如何决定我们的细胞寿命?
2025年11月10日 02:12

“我们为何会变老?”这个古老的问题,在微观的细胞世界中,有一个精准的分子答案正在浮现。它不在复杂的基因序列深处,而在每条染色体的末端——一段名为"端粒"的特殊结构。
伊丽莎白·布莱克本(Elizabeth H. Blackburn)因与卡罗尔·格雷德(Carol W. Greider)及杰克·绍斯塔克(Jack W. Szostak)因为共同发现端粒和端粒酶保护染色体的机制,获2009年诺贝尔生理学或医学奖。端粒位于染色体末端,像“帽子”一样防止基因组退化,维持细胞分裂稳定性。

伊丽莎白·布莱克本(Elizabeth H. Blackburn)、卡罗尔·格雷德(Carol W. Greider)、杰克·绍斯塔克(Jack W. Szostak)因为共同发现端粒和端粒酶保护染色体的机制,获2009年诺贝尔生理学或医学奖。
想象一下,我们的染色体如同一根长长的鞋带,而端粒就是鞋带两端包裹着的塑料帽。这个小小的“帽饰”至关重要:它防止鞋带(染色体)末端磨损、粘连,确保遗传信息能够被准确、完整地复制。每一次细胞分裂,每一次生命蓝图的拓印,这个保护帽都会不可避免地磨损、缩短一点。
当端粒缩短到一个临界长度,细胞便收到一个明确的信号:“生命时钟”即将鸣响。它不再分裂,进入一种名为“细胞衰老”的静默状态,或是走向程序性死亡。这不仅是细胞个体生命的终结,更是组织功能衰退、器官老化乃至个体衰老的微观起点。端粒的长度,因此被视为衡量细胞年龄和活力的“分子标尺”。
科学家们早已将“端粒损耗”列为国际公认的衰老核心标志之一。最新的研究正突破“关联”层面,逐步证实二者的因果关系:端粒不仅仅是衰老的被动记录者,更是驱动衰老进程的主动开关。理解端粒如何被磨损,又如何被守护,或许就是解开衰老之谜、干预衰老进程的关键所在。
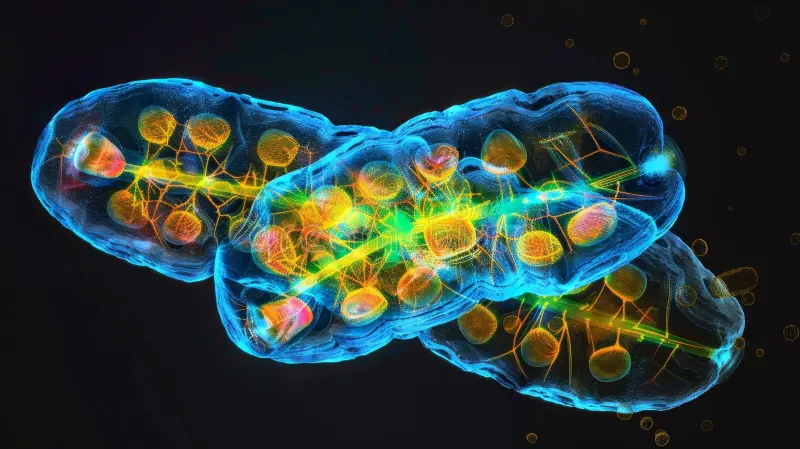

端粒是蛋白质-DNA复合物,保护染色体末端免受非法连接和切除。端粒酶是一种核糖核蛋白酶,合成端粒DNA以对抗端粒缩短。人类端粒是由端粒DNA和一种称为shelterin的六种蛋白质复合物组成的。它的主要功能可以概括为三点:保护、区分和计数。
保护功能是最直观的。通过端粒酶,TTAGGG重复序列阵列指定了人类染色体的末端。由6个端粒特异性蛋白组成的复合体与该序列相关,并保护染色体末端。线性染色体的末端容易被细胞内的DNA修复机制误认为是双链断裂损伤,从而引发不必要的修复反应,导致染色体末端融合、重排,引发基因组不稳定。端粒的独特结构和 Shelterin 蛋白复合体如同给染色体戴上了一顶“安全帽”,有效地"隐藏"了染色体末端,避免了这种灾难性的识别和修复。
区分功能在于,端粒明确了染色体DNA的“起点”和“终点”, 确保在DNA复制时遗传信息的完整性不被破坏。
最核心的功能是计数,即决定细胞的“海弗利克极限”。20世纪60年代,列奥纳德·海弗利克(Leonard Hayflick)发现,正常的人类体细胞在体外培养时,分裂次数存在一个上限,大约为40至60次,随后便会进入衰老状态。这一界限的分子基础,正是端粒的长度。
由于DNA复制机制固有的缺陷(即“末端复制难题”),染色体末端的端粒序列在每次细胞分裂后都无法被完全复制,从而导致端粒逐渐缩短。
初始的端粒长度如同一根“生命蜡烛”,燃烧(分裂)一次就短一截。当端粒耗尽后,细胞失去重要基因功能而衰老死亡。这一机制可能是多细胞生物衰老的根本原因之一。当它缩短到一个临界长度,细胞便会启动衰老或凋亡程序,以防止因基因组不稳定而可能导致的癌变风险。因此,端粒损耗是细胞一种重要的抗癌屏障。


端粒的自然缩短是不可避免的,但生活中的许多因素会极大地加速这一过程,如同有一只无形的手,在快速拨动我们的“生命时钟”。
1. 氧化应激:生化反应的“锈蚀”
细胞在代谢过程中会产生活性氧(ROS),这些高活性分子会攻击包括DNA在内的各种细胞组分。氧化应激是调节端粒丢失的重要因素,而端粒驱动的复制性衰老,本质上是细胞对端粒损耗应激的一种保护性应答。这一机制可能在进化中形成,以阻止那些暴露于高突变风险的细胞继续生长。慢性炎症、吸烟、空气污染(PM2.5)等都会显著增加体内的氧化应激水平,从而成为端粒的“头号杀手”。
2. 慢性炎症:持续燃烧的“暗火”
炎症是身体应对伤害和感染的防御机制,但当炎症变为慢性、低度持续时,它便从保护者变成了破坏者。持续的炎症状态会通过多种途径加速端粒磨损:一方面,炎症反应中大量产生的ROS直接损伤端粒DNA;另一方面,为了应对组织损伤,免疫细胞和周边组织细胞需要加速分裂增殖,这本身就会消耗端粒长度。一项针对大量人群的研究发现,体内炎症标志物(如C反应蛋白,CRP)水平越高的人,其白细胞端粒长度通常越短。
在人群中,端粒会随着年龄的增长而缩短,在所有已被检查的复制体细胞中,包括成纤维细胞和白细胞 。因此,端粒长度可以作为细胞生物学年龄或细胞进一步分裂潜力的生物标志物。
3. 慢性心理压力:情绪对染色体的“侵蚀”
“压力山大"不仅是一种感受,更有着深刻的生理学影响。著名的“照顾者研究” 发现,那些长期承受巨大精神压力、需要照顾慢性病子女的母亲,其免疫细胞的端粒长度显著短于年龄相仿但压力水平较低的女性。其机制在于,慢性压力导致皮质醇等压力激素长期处于高水平,这会增加全身的氧化应激,并可能直接抑制端粒酶的活性。情绪,正在以我们看不见的方式,雕刻着我们的细胞年龄。
4. 不良生活方式:日常的“磨损”积累
睡眠不足:睡眠是身体进行修复的关键时期。研究发现,睡眠时间短、质量差与更短的端粒长度相关。一晚彻夜不眠就足以导致端粒维护相关基因的表达发生变化。
缺乏运动与久坐:规律的中等强度运动被证明与较长的端粒长度相关。相反,久坐不动的生活方式则会加速端粒缩短。运动可能通过减轻氧化应激和炎症来发挥保护作用。
5.不健康的饮食
大量摄入加工食品、含糖饮料以及高升糖指数的食物,会促进炎症和氧化应激,从而加速端粒磨损。而富含抗氧化剂(如水果、蔬菜中的维生素C、E)、Omega-3脂肪酸(如深海鱼类)和优质蛋白的饮食模式,则有助于保护端粒。


面对端粒必然缩短的命运,细胞并非完全坐以待毙。它拥有一个强大的逆转工具——端粒酶。
端粒酶是一种特殊的逆转录酶,它能够以自身携带的RNA为模板,合成TTAGGG重复序列,并将其添加到缩短的染色体末端,从而补偿甚至延长端粒长度。这一发现荣获了2009年的诺贝尔生理学或医学奖。
在人体中,端粒酶在生殖细胞、干细胞和某些免疫细胞中活性较高,这确保了这些需要不断分裂的细胞群体能够维持其再生潜力。然而,在绝大多数体细胞中,端粒酶的活性在发育早期后就受到严格抑制。
这一抑制具有深刻的进化意义:它是一种强大的抗癌机制。如果所有细胞都能无限地激活端粒酶,修复端粒,那么它们就可能获得无限分裂的潜力,大大增加癌变的风险。事实上,大约85%-90%的癌细胞都找到了重新激活端粒酶表达的途径,从而实现了“永生”。因此,端粒酶是一把不折不扣的"双刃剑":在正常细胞中,它的缺失导致衰老;在癌细胞中,它的存在驱动永生。
这也为抗衰老研究带来了一个核心悖论和挑战:我们能否安全地、可控地在正常细胞中激活端粒酶,以延缓衰老,同时又不增加癌症风险?

尽管直接、系统地激活端粒酶的药物仍处于早期研究和严格的临床评估阶段,以避免潜在的致癌风险,但我们仍可以通过多种生活方式和营养干预,来减缓端粒的磨损速率,甚至可能通过提升特定细胞的端粒酶活性来维护端粒健康。
1. 营养支持:提供端粒维护的“原料”与“协作者”
· Omega-3脂肪酸:研究表明,补充Omega-3脂肪酸(如EPA和DHA)能够降低细胞中的氧化应激水平,并与白细胞端粒长度的减缓缩短相关。
· 维生素D:维生素D受体在基因组上分布广泛,参与调控众多基因表达。观察性研究发现,血液中维生素D水平较高与较长的端粒长度相关。其机制可能与其抗炎和免疫调节作用有关。
· 多种维生素与矿物质:
- B族维生素(叶酸/B9、B12):作为一碳单位代谢的关键辅因子,它们为DNA(包括端粒DNA)的合成和甲基化修复提供必要的原料。缺乏这些维生素会导致尿嘧啶错误掺入DNA,增加DNA不稳定性,可能加速端粒缩短。
- 锌和镁:锌是数百种酶的辅因子,包括那些参与DNA复制和修复的酶。镁同样在DNA稳定中扮演角色。研究表明,充足的锌和镁摄入与较长的端粒长度相关。
- 多酚类抗氧化剂:如白藜芦醇(存在于葡萄、蓝莓中)、姜黄素等,它们通过激活SIRT1等长寿相关蛋白,可能间接对端粒维护产生积极影响。SIRT1被证实可以与端粒结合,帮助维持其异染色质结构和稳定性。
2. 生活方式干预:激活内源性保护机制
· 规律且适度的运动:运动是公认的端粒保护因素。特别是高强度间歇训练(HIIT) 和耐力训练,已被随机对照试验证明能够显著提高白细胞中的端粒酶活性,并减缓端粒缩短。运动通过改善代谢健康、减少炎症和氧化应激来发挥作用。
· 压力管理与冥想:针对慢性压力对端粒的侵蚀,正念冥想、瑜伽等压力管理技术显示出保护作用。一项研究发现,长期冥想者其端粒酶活性显著高于对照组。这为"静心养性"提供了坚实的分子生物学的证据。
· 充足与高质量的睡眠:保证每晚7-9小时的高质量睡眠,为细胞修复(包括潜在的端粒维护)提供了关键窗口。建立规律的作息,避免熬夜,是对端粒最基础的呵护。
3. 新兴技术与药物探索(仍在研究阶段,前景与风险并存)
· TA-65:一种从黄芪植物中提取的环烯醚萜类化合物,被报道在商业产品中作为端粒酶激活剂销售。一些小规模初步研究显示它可能在一定程度上有选择性地延长最短的端粒,但其长期安全性、有效性和对癌症风险的真实影响,仍需大规模、长期的独立临床研究来证实。
· 基因疗法:在动物模型中,科学家尝试使用病毒载体将端粒酶基因导入特定细胞。例如,2012年《EMBO Molecular Medicine》上发表的一项里程碑研究显示,在成年小鼠中通过基因疗法激活端粒酶,可以显著延缓衰老,并延长其中老年寿命,且未增加癌症发生率。但这距离人类应用还有很长的路要走,其安全性和可控性仍是巨大挑战。


端粒,这个染色体末端的微小结构,以其精确的长度变化,为我们提供了一扇窥视细胞生命本质的窗口。它告诉我们,衰老并非一个模糊的、被时间推着走的过程,而是一个由分子事件驱动、可以被量化、甚至在一定程度上可以被干预的生物学程序。
我们的日常选择——吃什么、如何运动、怎样应对压力、能否安睡——都在不断地向我们的细胞发送信号,影响着端粒酶的活性和端粒磨损的速率。这些微小的日常决策,累积起来,便构成了我们细胞衰老的轨迹。
虽然我们尚无法拥有"长生不老"的魔法,但通过科学的生活方式,我们完全有能力将衰老的进程从"快进"调整为"慢放",让健康的寿命(健康预期寿命)最大限度地接近自然的寿命(预期寿命)。守护端粒,就是在细胞层面投资我们的未来健康。下一次,当你选择步行而非久坐,选择沙拉而非炸鸡,选择平心静气而非焦虑不安时,请记得,你不仅仅是在度过一天,更是在为你染色体末端的那些微小"时钟"赋能,为更长、更健康的生命篇章书写可能。
参考文献及出处:
[1] Smith EM, Pendlebury DF, Nandakumar J. Structural biology of telomeres and telomerase. Cell Mol Life Sci. 2020 Jan;77(1):61-79. doi: 10.1007/s00018-019-03369-x. Epub 2019 Nov 14. PMID: 31728577; PMCID: PMC6986361.
[2] van Steensel, B., Smogorzewska, A., & de Lange, T. (1998). TRF2 protects human telomeres from end-to-end fusions. Cell, 92(3), 401–413.
[3] Hayflick, L., & Moorhead, P. S. (1961). The serial cultivation of human diploid cell strains. Experimental Cell Research, 25, 585–621.
[4] Olovnikov, A. M. (1973). A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. Journal of Theoretical Biology, 41(1), 181–190.
[5] von Zglinicki, T. (2002). Oxidative stress shortens telomeres. Trends in Biochemical Sciences, 27(7), 339–344.
[6] Hou, L., et al. (2015). Air pollution exposure and telomere length in highly exposed subjects in Beijing, China: a repeated-measure study. Environment International, 85, 96-103.
[7] Epel, E. S., et al. (2004). Accelerated telomere shortening in response to life stress. Proceedings of the National Academy of Sciences, 101(49), 17312–17315.
[8] Liang, G., et al. (2011). Associations between rotational night shift work and telomere length in nurses. Occupational and Environmental Medicine, 68(6), 417-422.
[9] Werner, C. M., et al. (2019). Differential effects of endurance, interval, and resistance training on telomerase activity and telomere length in a randomized, controlled study. European Heart Journal, 40(1), 34–46.
[10] García-Calzón, S., et al. (2015). Dietary total antioxidant capacity is associated with leukocyte telomere length in a children and adolescent population. Clinical Nutrition, 34(4), 694-699.
[11] Greider, C. W., & Blackburn, E. H. (1985). Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell, 43(2), 405–413.
[12] Shay, J. W., & Bacchetti, S. (1997). A survey of telomerase activity in human cancer. European Journal of Cancer, 33(5), 787–791.
[13] Kiecolt-Glaser, J. K., et al. (2013). Omega-3 fatty acids, oxidative stress, and leukocyte telomere length: A randomized controlled trial. Brain, Behavior, and Immunity, 28, 16-24.
[14] Richards, J. B., et al. (2007). Higher serum vitamin D concentrations are associated with longer leukocyte telomere length in women. The American Journal of Clinical Nutrition, 86(5), 1420–1425.
[15] Paul, L. (2011). Diet, nutrition and telomere length. Journal of Nutritional Biochemistry, 22(10), 895-901.
[16] Dai, D. F., et al. (2010). Age-dependent cardiomyopathy in mitochondrial mutator mice is attenuated by overexpression of catalase targeted to mitochondria. Aging Cell, 9(4), 536-544. (相关讨论见文中关于矿物质与氧化应激/衰老部分)
[17] Palacios, J. A., et al. (2010). SIRT1 contributes to telomere maintenance and augments global homologous recombination. The Journal of Cell Biology, 191(7), 1299–1313.
[18] Jacobs, T. L., et al. (2011). Intensive meditation training, immune cell telomerase activity, and psychological mediators. Psychoneuroendocrinology, 36(5), 664–681.
[19] Harley, C. B., et al. (2011). A natural product telomerase activator as part of a health maintenance program. Rejuvenation Research, 14(1), 45-56.
[20] Bernardes de Jesus, B., et al. (2012). Telomerase gene therapy in adult and old mice delays aging and increases longevity without increasing cancer. EMBO Molecular Medicine, 4(8), 691–704.
本文作者为大健康领域资深媒体人,某平台合伙人。内容由山东大学齐鲁医学院教授、博士生导师郝爱军复核。Bioπ中国美肤科学传播平台发布本文只是为了更多的信息参考,不代表任何有倾向性的投资意见或市场暗示。

THE END





















京公网安备11010502058609号